Farben, Farben, Farben
Beschreibung: Pigmente - wie entstehen sie, Grundfarbfaktoren - was bewirken sie
Kategorie: Farben, Genetik, Vererbung & Co.
Link zu diesem Beitrag: Alles auswählen
[url=https://www.kaninchenwissen.de/kb/viewarticle?a=10&sid=25fde37670530e68cf424bdcc8c4259e]Knowledge Base - Farben, Farben, Farben[/url]
Copyright http://www.kaninchenwissen.de
Verwendung des Textes an anderer Stelle nur mit Genehmigung der Autoren.
Autor: reh und NiceDay
Die Pigmente
Als Pigmente (lat. pigmentum = Farbe, Farbstoff) werden im Allgemeinen jene Substanzen bezeichnet, die dem menschlichen Auge als gefärbt erscheinen. Es handelt sich dabei um eine Klasse chemischer Verbindungen, die verschiedene Funktionen erfüllen.
Melanine (gr. melas bzw. lat. melanus = schwarz) sind Pigmente, deren Farben von gelb, orange und rot bis zu braun und schwarz reichen. Meist handelt es sich um hochpolymere Verbindungen, die sich von der Aminosäure Tyrosin oder verwandten Stoffen ableiten und in den Pigmentzellen der Haut (Melanozyten) gebildet werden. Melanine findet man praktisch in allen Stämmen des Tierreichs. Melanin ist auch ein auffallender Bestandteil vieler Geschwulstbildungen, die als Melanome bezeichnet werden.
Die kugeligen Pigmentzellen der Haut, die Melanozyten, befinden sich nahe der Basis des Haarfollikels und haben unregelmäßige Verzweigungen und Verästelungen, die beim Pigmenttransport eine bedeutende Rolle spielen. Die Pigmentzellen werden in Form von Melanoblasten bereits in einem sehr frühen Stadium des sich entwickelnden Embryos gebildet, durchwandern noch während des embryonalen Wachstums die Lederhaut, gehen in die Oberhaut über, bilden mit den melaninaufnehmenden Hautzellen eine funktionelle Einheit und treten zum Teil in die sich im Aufbau befindliche Haarzwiebel ein. Der verbleibende Rest verweilt als Stammzellpopulation bzw. als Vorläuferzellen in den Grundschichten der Epidermis und kann, wenn die Haare im Zuge des Haarwechsels abgestoßen werden, wieder in die Haarzwiebel einwandern.
Pigmentzelle
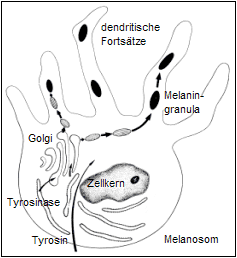
Das Enzym Tyrosinase wird im Golgi-Komplex angereichert. Die sich dort bildenden Melanosomen, eine Art Sekretbläschen, enthalten vor allem Tyrosinase und Melanin und stellen den eigentlichen Ort der Melaninsynthese dar. Die Melanosomen wandeln sich bei gleichzeitigem Verlust der Aktivität der Tyrosinase zu Melaninkörnchen um und wandern entlang der dendritischen Fortsätze in Richtung Spitze, wo sie konzentriert und in die umgebenden Zellen der Haut und in das Haar abgegeben werden.Die Verteilung der Melanosomen in den Pigmentzellen und deren Enzyminhalt (Tyrosinase) spielen bei der Pigmentbildung eine bedeutende Rolle.
Die Bildung der Fellfarbe wird bei Säugetieren im Wesentlichen von zwei Melaninen, nämlich Eumelanin und Phäomelanin, beeinflußt. In den Melanosomen wird aus dem Tyrosin mit Hilfe des Enzyms Tyrosinase sowohlEu- als auch Phäomelanin gebildet. Die Tyrosinase wirkt dabei als Katalysator, der in kleinsten Mengen einen chemischen Prozess (Melaninbildung) beschleunigt, ohne selbst in das Endprodukt einzugehen. An der Bildung von Phäomelanin ist außerdem noch das Eiweiß Cystein beteiligt.
Eumelanine sind ein wesentlicher Bestandteil der in braunen und schwarzen Haaren abgelagerten Pigmente und bilden dunkelbraune bis schwarze Pigmentkörner. Phäomelanine bewirken in Abhängigkeit von der Dichte ihrer Einlagerung gelbe bis rote Fellfarben. Je dichter die Einlagerung, desto intensiver die Farbe.
Um euch all zu viele Fachwörtern zu ersparen, bezeichne ich im Folgenden das Eumelanin als dunkles Pigment und das Phäomelanin als gelbes Pigment, die Melanozyten als Pigmentzellen.
Die meisten natürlichen Pigmente bestehen aus einer Basiskonzentration an gelbem Pigment und einer sich verändernden Konzentration an dunklem Pigment. Die Bildung von Kopolymeren bzw. Gemischen ist möglich. Ob nun dunkles oder gelbes Pigment gebildet wird, hängt neben der Aktivität der Tyrosinase vom Vorhandensein schwefelhaltiger Aminosäuren wie z. B. Cystein ab. Bei geringer Tyrosinaseaktivität wird bevorzugt gelbes Pigment gebildet. Die Aminosäure Cystein wird nicht nur bei der Synthese von gelbem Pigment, sondern auch beim Aufbau des Haares benötigt. Da das Kaninchen nicht in der Lage ist, Verbindungen wie Tyrosin und Cystein selbst herzustellen, müssen sie als essentielle (lebensnotwendige) Aminosäure über das Futter aufgenommen werde.
Die Bildung der Pigmente kann zeitlich begrenzt während des Haarwachstums erfolgen (ein Wechsel von dunklem zu gelbem Pigment ist möglich) oder regionale Unterschiede aufweisen. Die Pigmentzellen haben eine Kontaktstelle, an die sich verschiedene Eiweiße binden können (siehe Kasten unten), nämlich das Melanozyten-Stimulierende Hormon (MSH) und ein Eiweiß, welches mit dem Gen G zusammenhängt (siehe dort). Ich nenne es G-Eiweiß.
Zusammenfassung:
- Die Fellfarben der Kaninchen setzen sich aus dunklen und gelben (roten) Pigmenten zusammen, welche in speziellen Zellen gebildet werden.
- Die Pigmentzellen haben eine Kontaktstelle, an die sich entweder das MSH oder das G-Eiweiß binden kann.
- Bindet sich MSH an die Kontaktstelle, kommt es zu einer Erhöhung der Menge bzw. Aktivität der Tyrosinase und damit zur vermehrten Bildung von dunklem Pigment. Die Bildung von gelbem Pigment nimmt ab.
- Bindet sich das G-Eiweiß an die Kontaktstelle, kommt es zum gegenteiligen Effekt. Die Menge bzw. Aktivität der Tyrosinase nimmt ab, folglich wird vermehrt gelbes Pigment gebildet und die Bildung von dunklem Pigment nimmt ab.
Der Kampf um die Kontaktstelle
Ist die Kontaktstelle vom MSH belegt, wird vermehrt dunkler Farbstoff gebildet, ist sie vom G-Eiweiß (Agouti-Protein) besetzt, kommt es zur vermehrten Bildung von rotem Farbstoff. Die Bildung der Farbstoffe kann im Zuge des Haarwachstums zeitlich (Bänderung) und örtlich (Wildfarbigkeitsabzeichen) wechseln.

Wenn ich mich im weiteren Text auf dieses Bild beziehe, nenne ich den dunklen Herrn Eumel und den roten Phäo.Ist die Kontaktstelle vom MSH belegt, wird vermehrt dunkler Farbstoff gebildet, ist sie vom G-Eiweiß (Agouti-Protein) besetzt, kommt es zur vermehrten Bildung von rotem Farbstoff. Die Bildung der Farbstoffe kann im Zuge des Haarwachstums zeitlich (Bänderung) und örtlich (Wildfarbigkeitsabzeichen) wechseln.
Die symbolische Darstellung der Fellfarben-Faktoren
Es ist bereits seit langem bekannt, dass an der Entstehung der Haarfärbung des Kaninchens zahlreiche Erbanlagen beteiligt sind. Mehrere Hauptgene, welche unterschiedliche Merkmale kontrollieren, wurden identifiziert. Diese Hauptgene sind jedoch nicht die einzigen Gene, die ein Merkmal beeinflussen. Ein Organismus ist mit einer hoch komplexen Anlage vergleichbar, wobei alle Funktionen mehr oder weniger ineinandergreifen. Die Wirkung eines Gens auf die Ausbildung eines Merkmals erfolgt ebenfalls nicht völlig isoliert, sondern ist im Zusammenspiel zu betrachten. Die Ausprägung von Merkmalen hängt daher nicht nur von der Wirkung des betreffenden Gens ab, sondern auch von den Funktionen anderer Gene und der Umwelt.Erbfaktoren werden mit Hilfe von Buchstaben symbolisch dargestellt. Im deutschsprachigen Raum werden vor allem die von Prof. Dr. Hans Nachtsheim verwendeten Gensymbole benutzt. In der englischsprachigen Literatur werden diese Gene mit abweichenden Buchstaben bezeichnet. Die international gebräuchlichen Abkürzungen sind bei Kenntnis der entsprechenden Übersetzung zum Teil selbst erklärend und tragen dadurch zum besseren Verständnis bei.
Die Fellfarbe der Säugetiere ist ein Merkmal, das auf genetischer Ebene sehr gründlich untersucht wurde. Ein Großteil der beim Kaninchen bekannten Mutationen wurde bereits Anfang bis Mitte des 20. Jahrhunderts beschrieben. Bei diesen handelt es sich meist um funktionelle Störungen der Pigmentzellen, die vor allem deren Differenzierung, Vermehrung und Wanderung betreffen, oder um Störungen der beteiligten Enzyme, deren Aktivität eingeschränkt wird oder verloren geht. Bei der Beschreibung der Gene A, B, C, D und G wurde einerseits das Buch „Vom Wildtier zum Haustier“ von Prof. Dr. H. Nachtsheim zu Rate gezogen, andererseits auch Erkenntnisse aus dem Themenkreis „Maus“ (Jackson, 1994) verarbeitet.
Die Farbfaktoren A, ad , ach, am, an und a
Die Faktoren sind in der Reihenfolge ihrer absteigenden Dominanz geordnetDas Allel A . . . . . (int. C = color)
Der Faktor A ist als Grundfaktor der Pigmentbildung für die Entwicklung der Fellfarbe unerlässlich. Nur beim Vorhandensein dieses Faktors in rein- oder spalterbiger Form kann in Haut, Haar und Auge Pigment gebildet werden. International ist der Faktor A durch C ( color = Farbe) gekennzeichnet. Die Abkürzung spricht für sich. Es wurde bereits erwähnt, dass es sich beim Enzym Tyrosinase um das Schlüsselenzym der Melaninsynthese handelt. Wenn A die Pigmentbildung ermöglicht, dann muss dieses Gen für die Bildung der Tyrosinase zuständig sein.
Das Allel ad . . . . . (int. cchd)
ad wird als Faktor für Dunkelchinchilla bezeichnet. Tiere, die diesen Faktor tragen, sind etwas dunkler als chinchilla, da ein kleiner Teil des gelben Pigments noch gebildet werden kann; der Unterschied ist jedoch gering. Oft ist ein brauner Anflug in der Nestwolle zu finden (Zwischenfarbe). ad beschreibt nicht die in Würfen von Groß- und Kleinchinchilla vereinzelt auftretenden schwarzen Jungtiere. Deren Elterntiere sind möglicherweise hinsichtlich der Erbanlage G (Wildfarbigkeitsfaktor) spalterbig (Kombination G/g).
Das Allel achi . . . . . (int. cch)
Der Faktor achi - Chinchilla - verhindert die Bildung von gelbem Pigment und läßt sich daher als wildfarbig minus gelb definieren. Die zonale Gliederung des Haares bleibt erhalten, anstelle eines gelben tritt jedoch ein weißes Band in Erscheinung (achiBCDG). In Verbindung mit den Faktoren BCDgo führt achi zum Weißgrannen-Kaninchen. achi in Verbindung mit den Faktoren bCDG (gelb) ergibt ein fast weißes Tier, das nur noch pigmentierte Augen und Haarspitzen aufweist ( = Schwarzgrannen).
Die Ursache für das Fehlen von gelbem Pigment dürfte in einer verminderten Aktivität der Tyrosinase liegen. Die Bildung von dunklem Pigment ist nicht betroffen.
Das Allel am . . . . . (int. cchl)
Dem Faktor am, der als Marderfaktor bezeichnet wird, liegt ein stärkerer Abbau der Pigmente zugrunde. Als mögliche Ursache kommt ebenfalls eine verminderte Aktivität der Tyrosinase in Frage. Dunkles und gelbes Pigment werden im einzelnen Haar bzw. über das gesamte Fellkleid – den unterschiedlichen Aktivitätszuständen des Enzyms entsprechend – ungleichmäßig verteilt.
Standardgerechte Marderkaninchen tragen die Farbformel am/anBCDg. Es entsteht der Eindruck, als ob am für den Rückenstreifen bzw. die helle Seite verantwortlich ist; an dürfte federführend bei der Ausprägung der Abzeichen an Kopf, Lauf und Blume sein.
Das Allel an . . . . . (int. ch - himalayan)
Der Faktor an ist maßgeblich an der Bildung der als Russenzeichnung bekannten Fellfärbung beteiligt. Diese wird durch eine temperaturabhängige Mutante der Tyrosinase verursacht, die nur bei einem entsprechenden Kältereiz arbeitet. Nur die „äußeren“, kürzer behaarten Körperteile, die „Spitzen“ des Körpers sind pigmentiert. Das vor allem im Bereich der Extremitäten aktive Enzym führt zu dunklen Läufen, dunkler Blume, dunkler Nase und dunklen Ohren. Diese Erscheinung wird auch Akromelanismus oder Spitzenfärbung genannt. Im „warmen“ Bereich des Rumpfes ist das Enzym inaktiv und Haut, Haar und Auge sind pigmentlos. Die Jungtiere werden als Albinos geboren, die Abzeichen treten erst nach einigen Wochen in Erscheinung.
Das Allel a . . . . . (int. c)
Der Faktor a verhindert bei reinerbigem Auftreten die Pigmentbildung und damit das Sichtbarwerden anderer an der Ausprägung der Fellfarbe beteiligter Gene.
a/a-Tiere werden als Albinos (lat. albus = weiß), deren Fellfarbe als albinotisch-weiß bezeichnet. Haut, Haar und Auge sind pigmentlos. Die rote Farbe der Augen entsteht nicht durch Pigmente, sondern durch Reflektion des einfallenden Lichtes vom mit Adern durchzogenen Augenhintergrund. Albinos findet man beim Menschen, bei vielen Säugetierarten, gelegentlich sogar bei Vögeln, Amphibien und Fischen.
Da A als Gen für die Tyrosinase bezeichnet wurde, ist a mit Tyrosinase-negativ zu umschreiben. Aufgrund eines genetischen Defekts der Tyrosinase, welcher zum Verlust der Enzymaktivität führt, kann kein Pigment gebildet werden. Die Pigmentbildung unterbleibt, obwohl Melanozyten in normaler Zahl vorhanden sind.
Bei a handelt es sich um einen rezessiven Farbfaktor. Albinotische Tiere tragen verdeckt Farb- und/oder Zeichnungsanlagen. Diese treten nur bei der Verpaarung mit pigmentierten Tieren in Erscheinung. Mit überraschenden Ergebnissen ist zu rechnen. Beispielsweise können bei der Paarung Blauer Wiener x Weißer Neuseeländer neben hell- und dunkelgrauen auch blaue bzw. schwarze Tiere auftreten.
Die Farbfaktoren G, go und g
Die Faktoren sind in der Reihenfolge ihrer absteigenden Dominanz geordnetDas Allel G . . . . . (int. A = agouti)
G ist ein Faktor, der die Verteilung der Pigmente an den verschiedenen Körperteilen und im einzelnen Haar regelt. Meist wird die Erbanlage G Wildfarbigkeits- bzw. Farbverteilungsfaktor genannt. Im internationalen Sprachgebrauch wird das betreffende Gen bzw. der Phänotyp mit agouti bezeichnet. Die Gesamtfärbung ist kaum exakt beschreibbar – grau meliert trifft noch am ehesten zu – und entspricht dem Farbton des Wildkaninchens, welcher eine vorzügliche Anpassung an dessen Lebensraum darstellt.
Die Pigmente sind im Haar zonenweise bzw. ringförmig angeordnet. Bläuliche, schwarze, braune und gelbe Zonen wechseln einander ab. Die Bildung von gelbem und dunklem Pigment und deren Einlagerung ins Haar während des Haarwachstums erfolgt zu verschiedenen Zeitpunkten.
Eumel und Phäo sind ungefähr gleich stark, mal hat der eine die Oberhand, mal der andere, deshalb entstehen gestreifte Haare, an denen man den Verlauf des Kampfes erkennen kann.
Dunkles Pigment wird vor allem zu Beginn und am Ende des Wachstumszyklus gebildet, während gelbes Pigment hauptsächlich zur Mitte des Zyklus entsteht und als zentral angeordnetes gelbes Band bzw. gelber Streifen zu erkennen ist. Die Ausfärbung der jeweiligen Farbzonen wird durch die mengenmäßige Verteilung der Pigmentkörperchen bestimmt und führt zu einer entscheidenden Änderung der Gesamtfärbung des Felles. Fließende Übergänge von dunkel- bis hellgrau sind möglich.Wenn das vom Faktor G gebildete Eiweiß (Agouti-Protein) an der Kontaktstelle der Pigmentzellen sitzt, leitet diese den Wechsel der Bildung von dunklen zu gelben Pigmentkörperchen ein. Diese werden anschließend in das sich entwickelnde Haar eingebaut.
Die Aktivität dieses G-Eiweißes ist zeitlich begrenzt und demzufolge am einzelnen Haar als heller Streifen sichtbar. Besonders gut kann man diese Bänderung im sogenannten Haartrichter sehen, der sich bildet, wenn man dem Kaninchen ins Fell bläst.
Neben dem G-Eiweiß spielt das bereits genannte MSH eine wichtige Rolle. MSH kann ebenfalls an diese Kontaktstelle andocken und die Bildung von dunklem Pigment einleiten / anregen. Das Hormon und das G-Eiweiß konkurrieren um die Kontaktstelle. Welches Pigment gebildet wird, hängt sowohl vom jeweiligen Gegenspieler als auch vom Zustand der Kontaktstelle ab.
Neben der charakteristischen Bänderung des Haares weist die wildgraue Farbe weitere Eigenheiten auf. Der Österreichische Einheitsstandard beschreibt sie wie folgt:
Bei genauer Betrachtung ist jedes einzelne Deckhaar von unten nach oben gesehen blau, braun, gelb und schwarz. Das Blau entspricht der Unterfarbe und das Braun der Zwischenfarbe. Der sehr schmale gelbe und noch schmalere schwarze Ring verursacht eine fast undefinierbare graue Tönung. Über dieser Tönung ergeben die schwarzen Grannenspitzen, entsprechend ihrem Vorhandensein, eine helle, mittlere oder dunkle Schattierung. Die Brust und die Läufe sind meist etwas heller, der Kopf und die Ohren meist etwas dunkler als der Rücken. Der kleine Genickkeil ist bräunlich. Die Zwischenfarbe ist bräunlich bis rostbraun. Die Unterfarbe ist am ganzen Körper, einschließlich des Bauches, blau. Die Ohren sind kräftig schwarz umrandet. Die Blumenoberseite muss sehr deutlich eine Art grauer Perlung zeigen. Die Wildfarbigkeits(ab)zeichen sind deutlich ausgeprägt. Die Augen- und Kinnbackeneinfassung, die Einfassungen der Nasenöffnungen, die Bauchdeckfarbe, die Innenseite der Läufe und die Unterseite der Blume sind weiß.
Es ist offensichtlich, dass die Aktivität des G-Eiweißes während des Haarwachstums nicht nur zeitlich begrenzt erfolgt, sondern auch in den einzelnen Körperregionen unterschiedlich geregelt ist. Wie könnte sonst die unterschiedliche Verteilung gelber und dunkler Pigmente im einzelnen Haar bzw. über das gesamte Fellkleid erklärt werden. Am Rücken, mit Ausnahme des Genickkeils, werden dunkles und gelbes Pigment ungleich verteilt, wobei das G-Eiweiß als kurzer Puls agiert, der zu einer verminderten Aktivität der Tyrosinase führt. Gelbe Pigmentkörner werden folglich verstärkt eingelagert und sind als gelbes Band zu erkennen. Am Kopf ( Augen, Nase, Ohren) und am Bauch bzw. an den Läufen führt das G-Eiweiß zu einer verminderten bzw. kaum vorhandenen Einlagerung von Pigmenten. Die Fellfarbe der betreffenden Regionen ist weiß (Bauch) bis cremefarbig (Augen, Nase, Ohren). Die Bauchunterfarbe kann, muss aber nicht zwingend blau sein. Es gibt z.B. Castor-Rexe, die eine durchgehend weiße Bauchfarbe aufweisen. Bereiche wie die Schamgegend (Schoßflecken) bzw. die Umgebung der Saugwarzen können gelbes Pigment enthalten.
Das Allel go . . . . . (int. at = tan)
Die Erbanlage go wird nach wie vor in irreführender Weise als Lohfaktor bezeichnet. Treffender wäre die von Hochstrasser verwendete Bezeichnung Abzeichenbewahrungsfaktor, wobei ich persönlich eher für Teilagouti oder teilwildfarbig plädiere.
Die Namensgebung beruht wohl auf dem Umstand, dass der Faktor erstmals beim Lohkaninchen in Erscheinung trat. Das Weißgrannenkaninchen war zum damaligen Zeitpunkt nicht bekannt. Die nicht nur Lohkaninchen betreffende Funktion des Faktors go besteht vor allem in der Erhaltung der mit dem Faktor G in Verbindung stehenden hellen Bauchseite bzw. der bereits beim wildfarbigen Tier heller gefärbten Abzeichen an Nase, Ohren, Augen und Nacken. Die wildfarbige Decke hingegen wandelt sich in eine einfarbige um.
Lohfaktor
Eumel hat sich auf den Rücken des Kaninchens verzogen und Phäo auf den Bauch, sie gehen sich aus dem Weg.
Die Wechselwirkungen zwischen Kontaktstelle und G-Eiweiß sind auf die Bereiche Kopf, Brust, Bauch und Lauf beschränkt. Hier führt ein Aktivitätsschub (verstärkte Bildung des G-Eiweißes)zur Einlagerung heller Melaninkörner und infolgedessen zu cremefarbigen Abzeichen. Der Rücken ist aufgrund fehlender Aktivität des Faktors einfarbig.Eumel hat sich auf den Rücken des Kaninchens verzogen und Phäo auf den Bauch, sie gehen sich aus dem Weg.
Das Lohkaninchen stellt in seiner heutigen Form keinen ursprünglichen Mutationstyp dar, sondern ist eine Kombinationsrasse. Die lohgelbe Färbung der Bauchseite und der charakteristischen Abzeichen beruht nicht auf der Wirkung des Faktors go, sondern auf das gelbe Pigment verstärkenden Faktoren, sogenannten Rot- bzw. Gelbmodifizierer. Ohne diese Rotverstärker weisen die Tiere eine helle, ganz schwach gelbliche Bauchfarbe, ähnlich der eines G-Tieres, auf. Die ersten schwarzen Lohkaninchen waren eher creme- als lohfarbig.
Der Lohfaktor verhindert auch nicht die Einlagerung dunkler Pigmente in die Lohzeichnung. H. Schmitt konnte das Einwandern von dunklem Pigment in die Abzeichen der Lohkaninchen durch den Eisengrau-Faktor Be nachweisen. Der Genotyp ABeCDg/ABCDgo zeigt eine graubraune Bauchfarbe.
Ich sehe in dieser Färbung die Wirkung von starken Schwarzmodifizierern.
Da sich der Faktor go mit anderen Faktoren, z.B. B, C, und D kombinieren läßt, wurden im Laufe der Jahre neben schwarzen, auch braune blaue und fehfarbige Lohkaninchen gezüchtet. Die Weißgrannen tragen ebenfalls den Faktor go, wobei die rötliche Färbung der Abzeichen durch den Einfluss des Faktors achi unterdrückt wird.
Das Allel g . . . . . (int. a)
Einfarbigkeit g
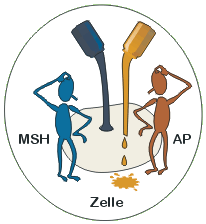
Eumel und Phäo können gar nichts tun, die dunkle Farbe ist fest mit der Kontaktstelle „verdrahtet“. (Das Beispiel hinkt, da Phäo in Verbindung mit gelb (b) doch zum Zuge kommt)
Der Faktor g führt zu einer relativ gleichmäßigen Verteilung der Pigmente im Haar. Die Bänderung des Haars unterbleibt. Die Farbintensität nimmt jedoch von der Haarspitze in Richtung Haarwurzel ab. Einfarbige Tiere besitzen neben einer dunklen Deckfarbe überwiegend eine hellere Unterfarbe. Während des Haarwachstums wird fortlaufend neues Pigment produziert, meist aber in verringertem Umfang, so dass die Unterwolle schwächer pigmentiert ist als die Decke. Das G-Eiweiß (das die Bildung von gelbem Pigment anregt bzw. einleitet) wird entweder nicht gebildet oder es ist funktionslos oder die Kontaktstelle ist so verändert, dass sie dauernd Signale für die Bildung von dunklem Pigment gibt. Ein Wechsel in der Bildung von dunklem zu gelbem Pigment findet nicht statt. Es wird vornehmlich dunkles Pigment in das wachsende Haar eingelagert.Eumel und Phäo können gar nichts tun, die dunkle Farbe ist fest mit der Kontaktstelle „verdrahtet“. (Das Beispiel hinkt, da Phäo in Verbindung mit gelb (b) doch zum Zuge kommt)
Die Farbfaktoren B, b, Be und bj
Die Faktoren sind an dieser Stelle nicht komplett in der Reihenfolge ihrer absteigenden Dominanz geordnet, bei den abweichenden Faktoren wird auf die Reihenfolge der Dominanz hingewiesen.Das Allel B . . . . . (int. E = Extension)
Der Faktor B wird meist als Faktor für dunkles bzw. schwarzes Pigment bezeichnet. Da Eumelanin ein wesentlicher Bestandteil schwarzer und brauner Haare ist, wird B auch Faktor für Eumelanin genannt. Im Englischen wird in diesem Zusammenhang der Begriff extension (= Ausdehnung, Ausbreitung des dunklen Pigments) verwendet. Faktor für die Schwarzausbreitung ist meiner Meinung nach eine sehr zutreffende Bezeichnung.
Eumel und Phäo sind ungefähr gleich stark, mal hat der eine die Oberhand, mal der andere, deshalb entstehen gestreifte Haare, an denen man den Verlauf des Kampfes erkennen kann
Das MSH regt die Bildung von dunklem Pigment an. Dies ist nur möglich, wenn die Kontaktstelle ihre volle Funktion besitzt. Besitzt sie diese nicht, werden dunkle Pigmente nicht oder nur zu einem geringen Teil produziert. Man kann folglich annehmen, dass die Gene B und b unterschiedliche Zustandsformen der Kontaktstelle darstellen.Das Allel b . . . . . (int. e)
Die meisten natürlichen Melanine bestehen aus einem Grundanteil an gelbem und einem veränderlichen Anteil an dunklem Pigment. Bei abnehmender Menge des dunklen Pigments müssen die Tiere zwangsläufig heller werden. Ein Übergang von schwarz zu braun und gelb tritt ein.
Das rezessive Allel b führt allerdings nur zu einem teilweisen Verlust an dunklem Pigment. Das läßt sich anhand des Thüringer-Kaninchens belegen, das durch die Farbformel AbCDg beschrieben wird. Trotz der Wirkungsweise des Faktors b und der durch den Faktor g bedingten gleichmäßigen Verteilung der Pigmente im Haar zeigen Thüringer keine reingelbe, sondern eine mit rußigem Schleier versehene Fellfarbe. In Verbindung mit dem Wildfarbigkeitsfaktor G entsteht mit AbCDG ein vom Thüringer-Gelb erheblich abweichendes Gelb. Als Beispiel ist das Burgunder-Kaninchen zu nennen. Wenn das G-Eiweiß die Bildung von gelbem Pigment anregt, müssen Tiere, die den Faktor in ihrem Erbgut aufweisen, heller bzw. gelber sein als jene, die ihn nicht tragen. Das Burgunder-Kaninchen z.B. ist eindeutig gelber bzw. roter als das Thüringer-Kaninchen.
Die Menge des übrig bleibenden dunklen Pigmentes, insbesondere des rußigen Anflugs, ist von der Stärke sogenannter Schwarzmodifizierer abhängig, wodurch dann Unterschiede wie zwischen den Schwarzgrannen und den Celler Weißen (wie Schwarzgrannen, aber ohne schwarze Grannenspitzen) entstehen. Deshalb kann der dunkle Anflug der Thüringer nicht durch die Kombination bg bedingt sein, denn er kann durch schwache Schwarzmodifizierer zurück gedrängt werden. Übrig bleiben dann dunkle Extremitäten, vergleichbar mit denen der Russenkaninchen.
Das Allel Be . . . . . (int. es =steel - Stahl)
Der Faktor Be, Faktor für Eisengrau, tritt am sichtbarsten in Kombination mit dem Wildfarbigkeitsfaktor G in Erscheinung und ist über B dominant. Die Tiere sind im gesamten sehr dunkel. Lediglich der Genickkeil und die Wildfarbigkeitsabzeichen sind etwas heller. Die Ohrenränder sind breit und kräftig schwarz eingefaßt; des öfteren sind die Ohren sogar gänzlich schwarz. Das Deckhaar ist in mehrere Zonen unterteilt. Die blaue Unterfarbe umfaßt etwa zwei Drittel der gesamten Haarlänge, die Zone der braunen Zwischenfarbe ist schmal und dunkel. Die bei wildgrauen Tieren noch gut sichtbare gelbe Zone ist kaum vorhanden. Die schwarzen Spitzen der Grannenhaare sind stark ausgeprägt, zum Teil sogar durchgehend schwarz gefärbt. Die Oberseite der Blume zeigt neben der schwarzen Grundfarbe eine leichte Durchsetzung mit einzelnen grauen Haaren, die eine Sprenkelung ergibt.
Be in Verbindung mit B (ABeCDG/ABCDG) führt aufgrund der verstärkten Produktion dunkler Pigmente zu einer Abnahme der Bänderung des einzelnen Haares. Dunkle Pigmentkörner wandern in die Wildfarbigkeitszeichen ein. Be vermindert bereits in spalterbiger Form die Wirkung des Faktors G, ist sozusagen eine Energiepille für Eumel. Es werden vermehrt dunkle Pigmentkörnchen gebildet, deren Einlagerung in das einzelne Haar zu einer einschneidenden Änderung der Gesamtfärbung des Felles führt. Dieser Phänotyp wird als Eisengrau bezeichnet, kann als wildfarbig mit verstärkter Einlagerung von dunklem Pigment beschrieben werden und tritt nur bei spalterbigen Tieren in Erscheinung.
- Be in reinerbiger Form (ABeCDg) verhindert die Zonenbildung des Haares und führt zu einem schwarzen Tier, das vom einfarbigen Schwarz der Alaskakaninchen (ABCDg) kaum zu unterscheiden ist. Be unterdrückt in reinerbiger Form die Wirkung des Faktors G nahezu vollständig. Ein Wechsel in der Melaninsynthese ist nicht möglich. Während des gesamten Haarwachstums wird vor allem dunkles Pigment gebildet. Ein Erkennen dieser Tiere ist nur an einzelnen etwas helleren Haaren (Leithaare) möglich.
- Be in Verbindung mit b (ABeCDG/AbCDG) ergibt schwarze Tiere (Manning et al.).
- Be in Verbindung mit bj (ABeCDG/Ab^je^ CDG) führt ebenfalls zu schwarzen Tieren (Schmitt).
Das Allel bj . . . . . (int. ej)
Die als Japanerfaktor bzw. Mosaikfaktor bekannte Erbanlage (international harlekin) führt zu einer mosaikförmigen Verteilung dunkler und gelber Pigmente und ist in der Dominanz zwischen B und b eingeordnet. Diese sind beidseits des Körpers als vertikale Streifen oder geblümte Felder unregelmäßig verteilt. Die einzelnen Farbfelder heben sich durch eine mehr oder weniger scharfe Abgrenzung voneinander ab. Am Bauch, an der Innenseite der Läufe und an der Blumenunterseite kann es durch G zu weißen Stellen kommen. In Bezug auf die Anordnung der Farbfelder ist eine große Variabilität möglich.
Der Einfluss des Faktors ist, ähnlich dem Lohfaktor, nicht auf die Rasse Japaner (AbjCDG ) beschränkt, sondern auch beim Rhönkaninchen (achibjCDG) sichtbar. Dort wird die gelbe Färbung der Farbfelder durch den Faktor achi verhindert.
Japaner-Kaninchen, die anstelle des Faktors G den Faktor g tragen, zeigen häufig schmutzig-gelbe Farbfelder, die dem Gelb des Thüringer-Kaninchens entsprechen. Allerdings unterliegt dieses Schmutziggelb in Bezug auf die Schwarzmodifizierer dem selben Mechanismus wie die Zeichnung beim Thüringerkaninchen und lässt sich vermutlich durch Selektion auf schwache Schwarzmodifizierer „wegzüchten“. Das Vorhandensein von G oder g kann man nur an den gelben Farbfeldern erkennen.
Die streifen- bzw. feldförmige Anordung der Pigmente ist auf wildfarbige bzw. lohfarbige Kaninchen übertragbar.
Bezüglich der Wirkungsweise des Japanerfaktors gibt es unterschiedliche Meinungen. In den letzten Jahrzehnten wurde von verschiedenen Autoren die Ansicht vertreten, dass es sich beim Faktor bj um eine Kombination der Faktoren Be und b handeln müsse. Be sorge für die tiefschwarzen, b für die gelben Farbfelder.
G. Hochstrasser vertritt den Standpunkt, dass der Faktor für die Verteilung der Farben in getrennte Farbfelder nicht an die B-Reihe gebunden ist, sondern außerhalb der Grundfarbformel steht. Er bezeichnet ihn als das selbständige, dominante Farbfelderteilungsgen J, welches als Modifikationsgen fungiert und die phänotypische Wirkung des Hauptgens quantitativ verändert. Seine ablehnende Haltung gegenüber der Kombination Be mit b begründet er unter anderem mit der Tatsache, dass Be in Verbindung mit b (die Verknüpfung zweier Allele sei nur in spalterbiger Form möglich - ABeCDG/AbCDG) schwarze Tiere ergibt.
Durch die Kontaktstelle auf den Pigmentzellen, die mit dem G-Eiweiß bzw. mit dem MSH arbeitet, kann der Faktor bj allerdings sehr gut unterschiedliche Zustandsformen dieser Kontaktstelle in verschiedenen Körperregionen darstellen und somit die Eigenschaften der Faktoren Be und b vereinen. Be sorgt für dunkle, b für gelbe Felder. Das würde bedeuten, dass die dunklen Farbfelder durch eine dauernd aktive Kontaktstelle und die gelben Felder durch eine teilweise funktionsunfähige Kontaktstelle verursacht würden. Der Faktor G spielt in Bezug auf die dunklen Felder keine Rolle, da das G-Eiweiß die Kontaktstelle nicht blockieren kann. Bezüglich der gelben Farbfelder ist eine Regulierung durch den Faktor G nur eingeschränkt möglich, da die Kontaktstelle lediglich zum Teil funktionsfähig ist. Eine grundlegende Funktion muss gegeben sein, da sich sonst Japanerkaninchen mit dem Faktor G von Japanern mit dem Faktor g bezüglich des Aussehens nicht unterscheiden würden. Sie tun es aber. Die Ausprägung der beiden Merkmale Be und bj erfolgt aufgrund der unterschiedlichen Zustandsformen der Pigmentzellen unabhängig voneinander. Der Japaner- bzw. Mosaikfaktor muss ein vorgeschalteter, übergeordneter Faktor sein, der bereits während der Embryonalentwicklung bei der Differenzierung der Melanoblasten zu Pigmentzellen und der Bildung der Kontaktstelle eine Rolle spielt.
Die Farbfaktoren C, c und D, d
Die Faktoren sind in der Reihenfolge ihrer absteigenden Dominanz geordnet.Das Allel C . . . . . (int. B)
Der Faktor C wird im Englischen mit brown bezeichnet. Im deutschen Sprachgebrauch wird C immer wieder mit schwarz gleichgesetzt und als Faktor für schwarze bzw. dunkle Pigmentierung beschrieben. Neuere Erkenntnisse weisen zumindest bei der Maus darauf hin, dass an der Melaninsynthese neben der Tyrosinase andere, mit dieser in Verbindung stehende, Proteine beteiligt sind, die einen Enzymkomplex bilden. Dunkles und gelbes Pigment können daher nur bei vollständiger Aktivität des Komplexes gebildet werden. W.E. Castle wies bereits 1924 auf eine Verbindung zwischen den Genen A (Tyrosinase) und C (schwarze bzw. braune Fellfarbe) hin.
Das Allel c . . . . . (int. B)
Der Faktor c wird meist als havannafarbig (englisch chocolate) bezeichnet. Aufgrund eines Aktivitätsverlustes eines der am Tyrosinase-Enzymkomplex beteiligten Enzyme (nicht der Tyrosinase!) nimmt die Pigmentierung des Haarkleides ab. Es erfolgt eine Aufhellung zu einem kräftigen Braun. Es ist anzunehmen, dass der Anteil an Eumelanin abnimmt. Da die Augen je nach Lichteinfall rot leuchten – die Blutgefäße im Augeninneren scheinen zum Teil durch – wurde die Rasse Havanna auch als „Feueraugen“ bezeichnet. Diese Erscheinung ist bei marder- und fehfarbigen Tieren ebenfalls beobachtbar. Der durch die Mutation von C zu c verursachte Pigmentverlust ist auch in den Krallen feststellbar.
Das Allel D . . . . . (int. D)
Der Faktor D bezeichnet die maximale Dichte der Pigmenteinlagerung. Er kontrolliert die Intensität der Pigmentierung, die durch die anderen für die Fellfarbe zuständigen Gene (ABCG) bestimmt wird. Im internationalen Sprachgebrauch wird D mit Dense (Dichte) oder Dilution (Verdünnung, Abschwächung, Milderung) gleichgesetzt.
Die Pigmentkörner in den Pigmentzellen werden in den Zellfortsätzen konzentriert und in die umgebenden Zellen von Haut und Haar abgegeben. Zur Anhäufung der Pigmentkörner in den Verästelungen ist ein Transportmechanismus erforderlich. Die maximale Dichte der Pigmenteinlagerung kann nur erreicht werden, wenn der Transport der Pigmentkörner reibungslos funktioniert.
Das Allel d . . . . . (int. d)
d führt zu einer verdünnten Einlagerung der Pigmente. Sowohl Eu- als auch Phäomelanin sind betroffen. Schwarz wird daher zu blau und gelb zu creme. Eine blaue Fellfarbe (Deck- und Unterfarbe) entsteht jedoch nur, wenn eine zonen- bzw. ringförmige Anordnung der Pigmente unterbleibt (siehe g) und eine gleichmäßige Verteilung erfolgt. Grund für die verringerte Pigmentdichte ist ein gestörter Transport. Die Pigmente werden dadurch verzögert und ungleichmäßig abgegeben. Die Pigmente selbst werden durch die Störung des Transportmechanismus nicht beeinflusst.